Die Sinnesorgane ermöglichen es, mit der Umwelt zu interagieren und Dinge außerhalb des Körpers wahrzunehmen. Zu den 5 Sinnen des Menschen gehören das Sehen, Tasten, Schmecken, Riechen und Hören/Gleichgewicht. Für die Sinneswahrnehmung braucht es einen adäquaten äußeren Reiz und eine spezialisierte Sinneszelle, die diesen Reiz wahrnehmen kann. Für die Weiterleitung und Verarbeitung der Reizinformationen sind zwei Prozesse von großer Bedeutung: die Transduktion zur Umwandlung des Reizes in ein Rezeptorpotenzial und die Transformation des Rezeptorpotenzials in ein Aktionspotenzial. Die entstandenen Aktionspotenziale werden zu spezifischen Hirnarealen geleitet für die Weiterverarbeitung der Informationen.
Kostenloser
Download
Lernleitfaden
Medizin ➜
Ein adäquater Reiz (mit minimalem Energieaufwand wird eine Reaktion des Sinnesorgans bewirkt) löst eine Erregung im Organ aus bspw. das Licht im Auge. Die Erregung wird über periphere Bahnen (bspw. Auge und N. opticus) weitergeleitet und in entsprechenden Hirnarealen (bspw. N. opticus und Corpus geniculatum laterale) verarbeitet. Es gibt also zwei zentrale Vorgänge:
Die vier wichtigsten klassischen Sinne sind das Sehen, Hören, Riechen und Schmecken. Ebenso gehört der Gleichgewichtssinn zu den Sinneswahrnehmungen.

Schematischer Aufbau des Auges
Bild: „Structure of the eye“ von Phil Schatz. Lizenz: CC0 1.0Um einen Gegenstand zu sehen, muss dieser auf dem Augenhintergrund abgebildet werden. Das Licht wird dazu am Übergang Luft-Augapfel gebrochen und das Bild des Gegenstandes auf dem Kopf stehend und verkleinert auf der Netzhaut Netzhaut Anatomie des Auges abgebildet. Die Gesamtbrechkraft des Auges beträgt 58,8 dpt (Cornea, Vorderkammer und fernakkomodierte Linse). Durch die Krümmung der Linse kann eine höhere Brechung erreicht werden und die Brechkraft somit an die Entfernung des Gegenstandes angepasst werden. Der Abstand zwischen Nah- und Fernpunkt, also dem minimalen und maximalen Abstand, bei dem eine scharfe Abbildung möglich ist, wird Akkomodationsbereich genannt. Den Unterschied zwischen den entsprechenden Dioptrien-Werten bildet die Akkomodationsbreite. Sie beträgt maximal 14 dpt. Die Pupille Pupille Physiologie und Anomalien der Pupille trägt zusätzlich zur Verschärfung des abgebildeten Gegenstands bei, indem sie störende Randstrahlen ausblendet.
Die Retina oder auch Netzhaut Netzhaut Anatomie des Auges kleidet den Augenbulbus aus. Im hinteren Bereich des Bulbus liegt die Pars optica, welche aus dem Stratum pigmentosum und Stratum nervosum besteht, im vorderen die Pars caeca, welche nur aus dem Stratum pigmentosum besteht. Von außen nach innen wird sie in folgende zehn Schichten gegliedert:
Stratum pigmentosum:
Stratum nervosum:
In der inneren Körnerschicht befinden sich zusätzlich Horizontalzellen, amakrine Zellen und Müller-Gliazellen, welche modulierend auf die Signalverarbeitung wirken und bspw. der Kontrastverstärkung dienen.
Es gibt zwei Arten an Photorezeptoren: Die Stäbchen vermitteln das Hell-Dunkel-Sehen oder auch skotopisches Sehen, die Zapfen das Farbsehen oder auch photopisches Sehen. Bei den Zapfen lassen sich drei Subtypen unterscheiden, die den drei Spektralfarben angepasst sind. Das menschliche Auge besitzt ca. 120 Mio. Stäbchen und etwa 6 Mio. Zapfen.
Der in den Sinneszellen enthaltene Sehfarbstoff 11-cis-Retinal absorbiert Licht und ändert seine Konformation zu All-trans-Retinal. Infolgedessen schließen die Natrium-Kanäle der Zellmembran Zellmembran Die Zelle: Zellmembran; es kommt zu einem hyperpolarisierenden Signal an den glutamatergen Photorezeptorsynapsen.

Retinale Isomere
Bild: „Retinal isomers“ von Phil Schatz. Lizenz: CC0 1.0Die Signale werden in einem Netz aus bipolaren Zellen, amakrinen Zellen und Horizontalzellen verarbeitet und auf Ganglienzellen verschaltet. Zapfen und Stäbchen sind demnach sekundäre Sinneszellen. In der Fovea centralis, dem Zentrum der Macula lutea, welche im Augenhintergrund auf der optischen Achse liegt, befinden sich nur Zapfen. Hier liegt der Ort des schärfsten Sehens. Medial der Macula lutea befindet sich die Papilla n. optici. Die Papilla n. optici kennzeichnet den Austritt des Nervus Opticus Nervus opticus Überblick über die Hirnnerven. Da aufgrund fehlender Photorezeptoren an dieser Stelle kein Sehen möglich ist, wird sie auch als blinder Fleck bezeichnet. Dieser liegt im Gesichtsfeld lateral.
Die Efferenzen der Ganglienzellen bilden den N. opticus. Dieser tritt aus dem Auge in der Papilla n. optici aus, verlässt die Orbita Orbita Orbita und extraokuläre Muskeln und tritt in die Schädelhöhle ein. Die Sehnerven beider Augen vereinigen sich im Chiasma opticum. Hier kreuzen die medialen Anteile der Netzhaut Netzhaut Anatomie des Auges auf die gegenüberliegende Seite, die temporalen Anteile bleiben ipsilateral. Daher verlaufen nach dem Chiasma opticum die ipsilateralen temporalen Fasern und die kontralateralen medialen Fasern gemeinsam. Der Tractus opticus projiziert dann zum Corpus geniculatum laterale im Thalamus Thalamus Thalamus. Von hier aus zieht die Sehbahn Sehbahn Die Sehbahn und Gesichtsfeldausfälle über die breit gefächerte Radiatio optica zur Sehrinde. Die Sehrinde gliedert sich in die primäre und sekundäre Sehrinde. Die primäre Sehrinde ist der Ort der Bewusstwerdung der visuellen Impulse, an die sich die Analyse der Informationen in der sekundären Sehrinde anschließt.
Aufgrund der Symptomatik können oftmals Rückschlüsse auf die Lage einer Läsion der Sehbahn Sehbahn Die Sehbahn und Gesichtsfeldausfälle geschlossen werden. Die folgenden Beispiele verdeutlichen dies:
Das Gehör verarbeitet Schallwellen – also Druckschwankungen in der Umgebung – zu elektrischen Signalen und nimmt somit Töne, Klänge und Geräusche wahr.
Die wichtigste Maßeinheit in diesem Zusammenhang ist der Schalldruckpegel. Er wird in Dezibel (dB) gemessen und beschreibt den Schalldruck in Relation zur Hörschwelle. Die Hörschwelle beschreibt den minimalen Schalldruck, bei dem ein Ton einer bestimmten Frequenz wahrgenommen wird. Durch die Relation zu einem bestimmten Wert (Hörschwelle) wird der Schalldruckpegel zu einer objektiv messbaren Größe. Er berechnet sich wie folgt: Schalldruckpegel = 20 x lg (p1 / p0). p0 ist der Bezugswert, also die Hörschwelle. Hiervon abzugrenzen ist der Lautstärkepegel. Er beschreibt die subjektive Empfindung der Lautstärke mithilfe der Phon-Skala. Die Phon-Werte entsprechen dem Schalldruckpegel bei einer Frequenz von 1 kHz. Das bedeutet z. B., dass Werte mit niedrigerer Frequenz einen höheren Schalldruckpegel benötigen als der Ton von 1 kHz, um genauso laut wahrgenommen zu werden. Diese Unterschiede werden durch Isophone verdeutlicht.
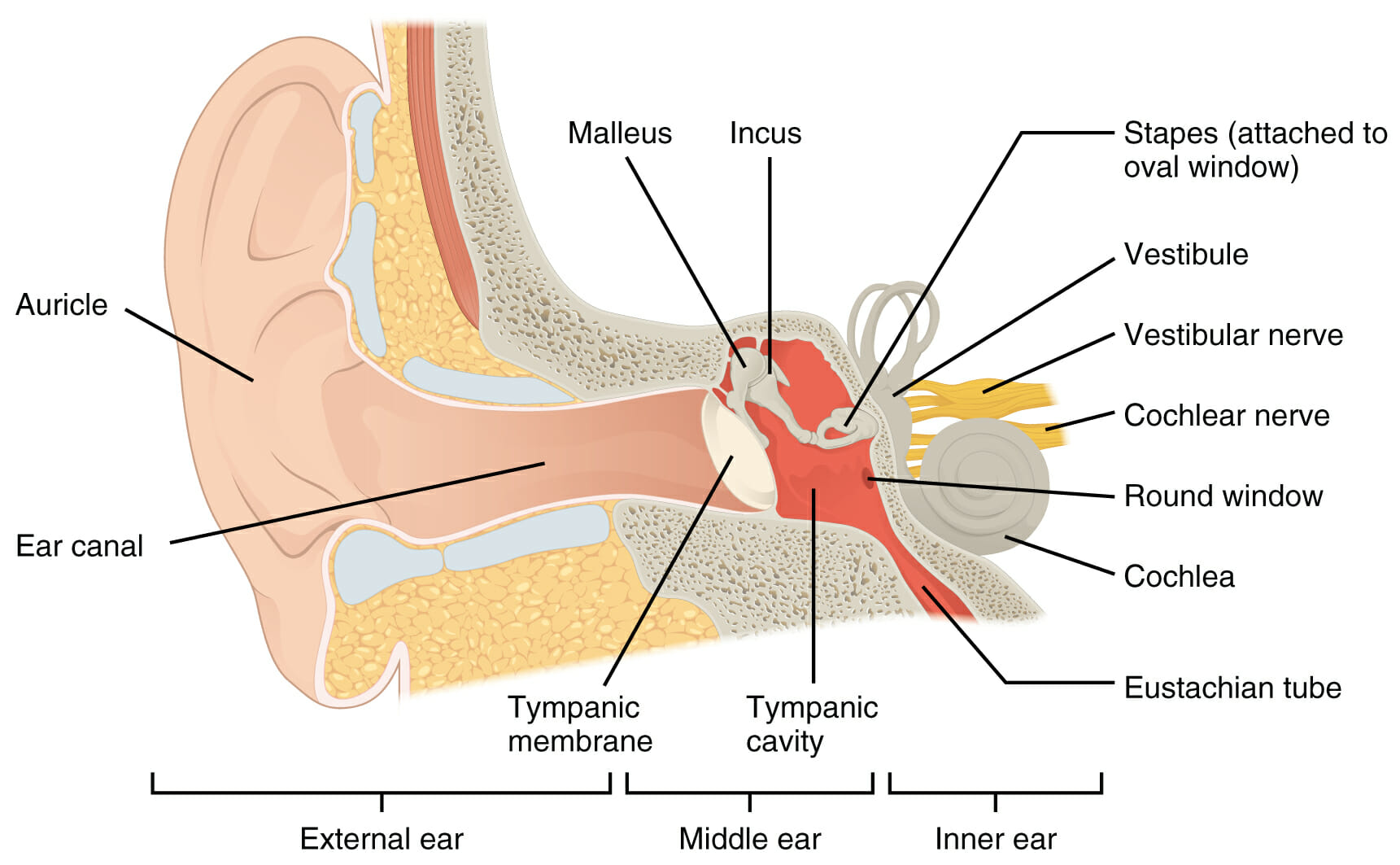
Schematischer Aufbau des Ohres
Bild: „Structure of the ear“ von Phil Schatz. Lizenz: CC0 1.0Das Ohr gliedert sich in das äußere Ohr, Mittelohr und Innenohr Innenohr Anatomie des Ohrs. Der Schall gelangt durch das äußere Ohr zum Trommelfell Trommelfell Anatomie des Ohrs. Dort wird er über die Gehörknöchelchen – Hammer, Amboss und Steigbügel – zum ovalen Fenster des Innenohrs geleitet. Im Mittelohr erfolgt die Impedanzanpassung, die eine Schallübertragung von 60 % ermöglicht. Ohne diese Anpassung würden 98 % des Schalles reflektiert werden. Die Anpassung erfolgt durch drei Mechanismen:

Aufbau der Cochlea
Bild: „1406 Cochlea“ von OpenStax. Lizenz: CC BY 4.0Der für das Hören verantwortliche Teil des Innenohrs ist die Cochlea. Die Cochlea besteht aus mehreren Schläuchen (der Scala tympani, Scala media und Scala vestibuli) und dem Corti-Organ, die zu der Hörschnecke aufgerollt sind. Die Scala tympani und Scala vestibuli enthalten die Natrium-reiche Perilymphe, die Scala media enthält die Kalium-reiche Endolymphe. Sie wird von der Reissner-Membran zur Scala vestibuli hin, von der Basilarmembran zur Scala tympani und von der Stria vascularis begrenzt. Das Corti-Organ enthält die eigentlichen Sinneszellen, die äußeren und inneren Haarzellen und wird von der Tektorialmembran bedeckt. Es gibt drei Reihen äußerer Haarzellen, welche mit ihren längsten Stereozilien die Tektorialmembran berühren und eine Reihe innerer Haarzellen, deren Stereozilien die Membran nicht berühren, sondern durch eine Verschiebung der Endolymphe (hydrodynamische Kopplung) ausgelenkt und somit erregt werden.
Die durch den Steigbügel ausgelöste Wanderwelle in der Scala vestibuli bewirkt eine frequenzspezifische, maximale Auslenkung der Basilarmembran an dem entsprechenden Punkt der Cochlea. Sie führt zu einer Scherbewegung der Tektorialmembran gegen das Corti-Organ und die Haarzellen. Hohe Töne werden nahe des ovalen Fensters abgebildet, tiefe Töne in Richtung des Helicotremas. Durch eine Auslenkung der Stereozilien entsteht ein Rezeptorpotenzial, eine mechanoelektrische Transduktion findet statt. Unerlässlich für die Potenzialbildung ist das endokochleäre Potenzial, welches durch die höhere Kalium-Konzentration der Endolymphe im Vergleich zur Perilymphe erzeugt wird. Aufgebaut wird dieses Potenzial durch die Stria vascularis, die aktiv Kalium-Ionen in die Endolymphe transportiert. Durch die Auslenkung der Stereozilien öffnen Kalium-Kanäle, Kalium aus der Endolymphe strömt ein und depolarisiert die Zelle. Anschließend öffnen Calcium-Kanäle und der Calcium-Strom resultiert in einer glutamatergen synaptischen Übertragung des Potenzials auf das erste Neuron der Hörbahn.
Das Perikaryon Perikaryon Nervensystem: Histologie des ersten Neurons befindet sich im Ganglion spirale. Diese bipolaren Zellen leiten die Potenziale weiter in den Ncll. cochleares der Medulla oblongata Medulla Oblongata Hirnstamm. Ihre Fortsätze bilden dabei einen Teil des N. vestibulocochlearis. Die tonotopische Gliederung der Schneckenwindungen bleibt erhalten. Nun kreuzt ein Teil der Fasern im Corpus trapezoideum, während ein Teil ungekreuzt weiterläuft. Der gekreuzte Teil wird u. a. in die Ncll. olivares superiores verschaltet, was essentiell für das Richtungshören ist. Kontralateral führen nun beide Stränge gemeinsam als Lemniscus lateralis zu den Colliculi inferiores. Unter Abgabe einiger weiterer Nebenäste und Rückkreuzungen zieht der Hauptteil der Fasern zum Corpus geniculum mediale und von hier als Hörstrahlung weiter zur primären Hörrinde. Die Hörrinde erhält somit Informationen aus beiden Cochleae. Diese wirkt sich positiv bei einer einseitigen Schädigung der Hörbahn und beim Richtungshören aus. In der primären Hörrinde erfolgt die Bewusstwerdung von Tönen. Die sinnvolle Verknüpfung zu Wörtern oder Melodien findet in der sekundären Hörrinde statt.
Neben der Cochlea befindet sich im Innenohr Innenohr Anatomie des Ohrs das Vestibularorgan, das für den Gleichgewichtssinn verantwortlich ist. Es setzt sich zusammen aus drei Bogengangsorganen und zwei Maculaorganen, die zusammen die Raumorientierung ermöglichen und Bewegungen registrieren.
Der Aufbau ähnelt der Cochlea. Die Schläuche des Vestibularorgans sind ebenfalls mit Endolymphen gefüllt. Die Sinneszellen sind ebenfalls Haarzellen. Sie bilden im Gegensatz zu cochleären Haarzellen ein Kinozilium und mehrere Stereozilien aus, die durch tip-links verbunden sind. Bedeckt werden sie von einer gallertartigen Masse. In den Bogengängen wird diese mukopolysaccharidhaltige Masse Cupula genannt. In den Maculaorganen enthält sie zudem kleine Kalziumkarbonatkristalle und wird deshalb Otolithenmembran genannt.
Durch das Beschleunigen, Bremsen oder Drehen des Kopfes wird die zähe Cupula/Otolithenmembran gegen die Sinneszellen verschoben. Es kommt wie in der Cochlea zu einer Scherbewegung, zu einer Auslenkung der Kinozilien sowie Stereozilien und dem Erzeugen eines Rezeptorpotenzials (siehe Abschnitt Hören). Der Transduktionsvorgang läuft in den Bogengängen und der Macula gleich ab; aufgrund ihrer anatomischen Unterschiede messen die Organe jedoch verschiedene Bewegungen. Translationsbewegungen: Das Beschleunigen und Bremsen wird von den Maculaorganen gemessen. Macula sacculi misst in senkrechter Richtung, Macula utriculi in waagerechter Richtung.

Lineare Bewegung im Gleichgewichtssystem
Bild: „Linear movement of the head ku“ von Phil Schatz. Lizenz: CC BY 4.0Rotationsbewegungen: Die Endolymphe der Bogengänge ist kreisförmig angeordnet. Bei der Drehung wird die Flüssigkeit aufgrund ihrer Trägheit gegen das Sinnesepithel des Bogengangs verschoben und die Zilien der Zellen ausgelenkt. Die Zilien sind so aufgebaut, dass sie bei einer Auslenkung nach medial hin zum Utrikulus ein Potenzial auslösen. Das heißt, bei einer Kopfdrehung nach links erfolgt eine Scherbewegung der Flüssigkeit in den horizontalen Bogengängen nach rechts, was in einer Aktivität des linken Bogenganges und den afferenten Nerven resultiert.

Gleichgewichtssinn
Bild: „1410 Equilibrium and Semicircular Canals“ von Phil Schatz. Lizenz: CC BY 4.0Die erzeugten Potenziale werden vom ersten Neuron als Teil des N. vestibulocochlearis auf den Ncll. vestibulares im Rhombencephalon Rhombencephalon Entwicklung des ZNS, Neurocraniums und Viscerocraniums und das zweite Neuron übertragen. Von hier zieht die Bahn gekreuzt und ungekreuzt in den Ncl. ventralis posterior des Thalamus Thalamus Thalamus. Anschließend werden die Impulse an die vestibulären Rindenfelder des Großhirns weitergeleitet.
Die Informationen des Vestibularorgans werden ständig verrechnet mit somatosensorischen Informationen aus dem Kopf und Halsbereich sowie aus anderen Gelenken, sodass das ZNS Informationen über die Gesamtkörperhaltung besitzt. Die beteiligten vier Vestibulariskerne sind Ncl. superior Bechterew, Ncl. inferior Roller, Ncl. medialis Schwalbe und Ncl. lateralis Deiters. Dies ermöglicht auch Muskelreflexe, die zur Erhaltung des Körpergleichgewichts ausgelöst werden. Besonders interessant sind die vestibulookulären Reflexe, welche das Vestibularorgan mit den Augenmuskeln verknüpfen. Dies spielt z. B bei einer Drehbewegung eine Rolle. Ein vestibulärer Nystagmus ist eine zusammengesetzte Augenbewegung, die aus einer langsamen, ausgleichenden Bewegung (vestibulär) und einer schnellen Rückstellbewegung besteht.
Beispiel: Wird eine Person auf einem Stuhl sitzend nach rechts gedreht, werden die Sinneszellen im rechten Bogenorgan aktiviert. Sie projizieren über die Vestibulariskerne zu den Augenmuskelkernen und bewirken eine Drehung der Augen nach links. Es folgt eine Blickstabilisation. Die schnelle Rückstellbewegung wird zentral vermittelt und folgt der Drehbewegung.
Dieser geruchswahrnehmende Bereich (die Regio olfactoria) ist beim Menschen stark zurückgebildet und bedeckt nur die obere Nasenmuschel sowie die gegenüberliegende Nasenscheidewand Nasenscheidewand Anatomie der Nase. Das mehrreihige Riechepithel besteht aus drei Zellpopulationen.
Die Basalzellen sind adulte Stammzellen und dienen der Erneuerung der nur ca. 30 Tage alten Riechzellen. Die Riechsinneszellen sind die primären, bipolaren Sinneszellen des Geruchsorgans. Sie bilden lange Kinozilien (Riechhärchen oder olfaktorische Zilien) aus, welche mithilfe von Chemorezeptoren Moleküle der Atemluft binden und somit die Sinneszelle erregen. Jede Sinneszelle kann nur eine Geruchsqualität wahrnehmen, wobei eine Geruchsqualität von Zehntausenden von Sinneszellen wahrgenommen wird. Der Mensch besitzt ca. 350 verschiedene Rezeptoren Rezeptoren Rezeptoren und kann sieben typische Geruchsklassen unterscheiden. Primäre Sinneszellen leiten die Erregung ohne eine Verschaltung über ihre Axone direkt in das ZNS weiter. Die Axone oder auch Fila olfactoria werden im N. olfactorius gebündelt und führen durch die Lamina cribrosa der Siebbeinplatte direkt in den Bulbus olfactorius, der als vorgelagerter Teil des ZNS zu betrachten ist. Hier findet die erste Verschaltung statt.
Im Bulbus olfactorius bilden die Endigungen der Fila olfactoria mit den Dendriten der Mitralzellen Glomeruli, welche die kleinsten funktionellen Einheiten des Geruchsorgans bilden. Dort befindet sich die erste Synapse Synapse Synapsen und Neurotransmission des Geruchsorgans. Dabei kommt es zu einer Konvergenz: Über tausend Axone der Sinneszellen projizieren auf die Dendriten einer Mitralzelle. Das Signal kann durch periglomeruläre Zellen und Körnerzellen in den Glomeruli moduliert werden. Durch eine laterale Hemmung können die periglomerulären Zellen das Signal des erregten Glomerulus verstärken und von benachbarten, schwächeren Signalen schärfer abgrenzen.
Die ca. 30.000 Axone der Mitralzellen verlassen den Bulbus als Tractus olfactorius. Er teilt sich in einen Hauptast und einen Nebenast. Der Hauptast kreuzt in der vorderen Kommissur zum Bulbus der gegenüberliegenden Hirnseite, wohingegen der Nebenast zum Riechhirn projiziert. Das Riechhirn befindet sich im Paleokortex und setzt sich aus vielen olfaktorischen Projektionsfeldern zusammen. Von hier werden die Informationen sowohl zum Neocortex und dem Cortex praepiriformis als auch zum limbischen System und von dort zu Kerngebieten des Hypothalamus Hypothalamus Hypothalamus und der Formatio reticularis geleitet.

Geruchssinn
Bild: „1403 Olfaction“ von Phil Schatz. Lizenz: CC BY 4.0Geschmack wird mit der Zunge Zunge Mundhöhle: Lippen und Zunge wahrgenommen, auf der sich die entsprechenden Strukturen befinden. Es gibt drei verschiedene Geschmackspapillen:
In den Geschmackspapillen finden sich die Geschmacksknospen. Es gibt 2.000–4.000 Knospen mit je 10–50 Sinneszellen. Die Knospen bilden einen mit Flüssigkeit gefüllten Porus aus, in den die Sinneszellen mit ihren Mikrovilli hineinragen. Auf den Mikrovilli befinden sich die eigentlichen Geschmacksrezeptoren.

Schematischer Aufbau einer Geschmacksknospe
Bild: „ Schematic drawing of a taste bud labelled in german“ von Jonas Töle. Lizenz: CC0 1.0Hier wird der chemische Reiz in ein elektrisches Signal umgewandelt. Die chemischen Stoffe/Verhältnisse lösen durch verschiedene Rezeptoren Rezeptoren Rezeptoren oder Kanäle immer eine Depolarisation der Sinneszelle aus, was zu einer Transmitterfreisetzung und Aktivierung innervierender Nerven führt. Die Geschmackssinneszellen sind somit sekundäre Sinneszellen. Es gibt vier verschiedene Geschmacksqualitäten, die wahrgenommen werden können: süß, sauer, salzig, bitter. Eine Sinneszelle kann entweder nur eine Geschmacksqualität erfassen oder alle vier, allerdings mit einer festgelegten Rangordnung für die vier Qualitäten.
Innerviert werden die Papillen vom N. glossopharyngeus und vom N. vagus. Diese Nerven ziehen in den Ncll. tractus solitarii des Hirnstamms. Die Information wird auf das zweite Neuron umgeschaltet und ipsilateral in den Ncl. parabrachialis der Formatio Reticularis auf das dritte Neuron weitergeleitet. Dieses Neuron projiziert in den kontralateralen Ncl. ventralis posterior des Thalamus Thalamus Thalamus. Vom Thalamus Thalamus Thalamus aus werden die Informationen in verschiedene Rindenbereiche übertragen. Hier erfolgen das Bewusstwerden des Geschmacks und die Verknüpfung mit anderen Wahrnehmungen, wie z. B. dem Geruch.



















